Thorpe et al (2007) Science, 316:1328-1331.
 A
fundamental question in human evolution is how modern humans and their
ancestors (hominins) became terrestrial bipeds. Human bipedalism is
traditionally thought to have evolved from a quadrupedal terrestrial precursor,
much like the common chimpanzee. But some recent paleontological and genetic
evidence suggests that adaptations for bipedalism must have arisen in an
arboreal context. However, the adaptive benefit of arboreal bipedalism has been
unknown. In this paper we hypothesised that bipedal locomotion might be
advantageous for arboreal apes moving on the thin terminal branches at the
periphery of trees because their long prehensile toes can grip multiple small
branches and thus maximize stability, while freeing one or both hands for
balance, feeding or weight transfer while crossing gaps between tree crowns. Our
results showed that bipedalism does indeed allow the most arboreal great ape,
the orangutan, to access supports too flexible to be negotiated otherwise.
Orangutans react to branch flexibility like humans running on springy tracks, by
straightening their knee and hip joints, whereas all other primates do the
reverse. We concluded that human bipedalism is less an innovation than an
exploitation of a locomotor behavior retained from the common great ape
ancestor.
A
fundamental question in human evolution is how modern humans and their
ancestors (hominins) became terrestrial bipeds. Human bipedalism is
traditionally thought to have evolved from a quadrupedal terrestrial precursor,
much like the common chimpanzee. But some recent paleontological and genetic
evidence suggests that adaptations for bipedalism must have arisen in an
arboreal context. However, the adaptive benefit of arboreal bipedalism has been
unknown. In this paper we hypothesised that bipedal locomotion might be
advantageous for arboreal apes moving on the thin terminal branches at the
periphery of trees because their long prehensile toes can grip multiple small
branches and thus maximize stability, while freeing one or both hands for
balance, feeding or weight transfer while crossing gaps between tree crowns. Our
results showed that bipedalism does indeed allow the most arboreal great ape,
the orangutan, to access supports too flexible to be negotiated otherwise.
Orangutans react to branch flexibility like humans running on springy tracks, by
straightening their knee and hip joints, whereas all other primates do the
reverse. We concluded that human bipedalism is less an innovation than an
exploitation of a locomotor behavior retained from the common great ape
ancestor.
for
technical comments on this paper and our response see Begun et al (2007) Science
318:1065 and Crompton and Thorpe (2007) Science
318:1065.
Thorpe et al. (2007) Biology Letters 3: 253-256
 Within
the canopy of tropical rainforest, the shortest gaps between tree crowns lie
between slender terminal branches. While the flexibility (compliance) of these
supports has previously been shown to increase the energetic cost of gap
crossing in arboreal animals, field observations suggest that some primates may
be able to use support compliance to increase the energetic efficiency of
locomotion. In this paper we calculated the energetic cost of alternative
methods of gap crossing in orangutans (Pongo abelii). Tree sway (see movie) was
found to be less than half as costly as jumping, and an order of magnitude less
costly than descending the tree, walking to the vine and climbing it.
Observations of wild orangutans suggest that they actually use support
compliance in many aspects of their locomotor behaviour. This study seems to be
the first to show that elastic compliance in arboreal supports can be used to
reduce the energetic cost of gap crossing.
Within
the canopy of tropical rainforest, the shortest gaps between tree crowns lie
between slender terminal branches. While the flexibility (compliance) of these
supports has previously been shown to increase the energetic cost of gap
crossing in arboreal animals, field observations suggest that some primates may
be able to use support compliance to increase the energetic efficiency of
locomotion. In this paper we calculated the energetic cost of alternative
methods of gap crossing in orangutans (Pongo abelii). Tree sway (see movie) was
found to be less than half as costly as jumping, and an order of magnitude less
costly than descending the tree, walking to the vine and climbing it.
Observations of wild orangutans suggest that they actually use support
compliance in many aspects of their locomotor behaviour. This study seems to be
the first to show that elastic compliance in arboreal supports can be used to
reduce the energetic cost of gap crossing.
O’Conner et al. (2007) Gait and Posture 25:469-474.
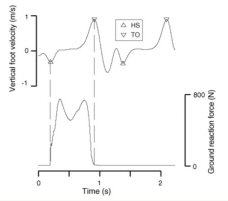 |
|
The vertical velocity of the foot centre of a man walking, with triangular markers indicating the timing of heel strike (HS) and toe off (TO) as determined by the foot velocity algorithm, and a simultaneous recording from a force plate which provides the ‘‘true’’ times for HS and TO for one step, which are marked with dashed lines. |
This study uses electromyography to establish the development of normal
activation patterns in the major muscles of the human leg during ontogeny. The
project, in collaboration with the University of Cape Town and University
College Dublin, will aid understanding of gait-related developmental problems,
such as cerebral palsy.
We are in the process of writing up this work, but have completed the first step
(!). The timing of heel strike (HS) and toe off (TO) are the events that mark
the transitions between stance and swing phase of gait and are important
measures when analysing gait. The timing of these events are normally identified
from kinetic (force) data, but these approaches restrict the number of steps
that can be analyzed. We developed an algorithm to automatically determine HS
and TO from kinematic (movement) data recorded by a motion capture system. The
foot velocity algorithm uses data from the heel and toe markers and
identifies features in the vertical velocity of the foot which correspond to the
gait events. Our foot velocity algorithm offers accurate results and is easy to
implement, and should be applicable in a variety of gait analysis settings.
 The
apes (including humans) share a suite of morphological adaptations in the trunk
and upper limb that have been interpreted by most as characteristics linked to
brachiation: orthograde (vertical trunk) forelimb-suspensory under-branch
locomotion. The Asian apes, more than any other, remain restricted to an
arboreal habitat and they are consequently an important model in the
interpretation of the morphological commonalities of the apes.
The
apes (including humans) share a suite of morphological adaptations in the trunk
and upper limb that have been interpreted by most as characteristics linked to
brachiation: orthograde (vertical trunk) forelimb-suspensory under-branch
locomotion. The Asian apes, more than any other, remain restricted to an
arboreal habitat and they are consequently an important model in the
interpretation of the morphological commonalities of the apes.
In this study we carried out a detailed analysis of orangutan positional
behavior (locomotion and posture). We show that orangutan positional behavior is
highly complex, representing a diverse spectrum of positional modes. Overall,
our results suggest that orangutans use orthograde suspensory behaviour
substantially less, but orthograde compressive locomotor modes (such as
hand-assisted bipedalism) far more than has been previously suggested. In
general orangutan positional behavior is similar to that of the African apes,
and in particular, lowland gorillas. In our conclusion we argue that it is
orthogrady in general, rather than forelimb suspension specifically, that is the
common inheritance from the last common-ape ancestor over 20 million years ago.
 The large body mass and exclusively arboreal lifestyle of Sumatran orangutans
identify them as a key species in understanding the dynamic between primates and
their environment. In this study we employ a multivariate modeling approach to
identify key associations between orangutan locomotion, canopy level, support
use and contextual behavior. Log-linear modelling is particularly appropriate
because it is designed for categorical data (such as is collected during
observational field studies), provides a systematic method for testing
alternative hypotheses regarding interactions between variables, and allows
interactions to be ranked numerically in terms of relative importance. The
diameter, type and number of supports used were found to have the strongest
associations with locomotor repertoire, suggesting that these variables, rather
than height, contextual behaviour, or the age-sex category of the orangutan,
have been the most important influences on the evolution of orangutan locomotor
profiles.
The large body mass and exclusively arboreal lifestyle of Sumatran orangutans
identify them as a key species in understanding the dynamic between primates and
their environment. In this study we employ a multivariate modeling approach to
identify key associations between orangutan locomotion, canopy level, support
use and contextual behavior. Log-linear modelling is particularly appropriate
because it is designed for categorical data (such as is collected during
observational field studies), provides a systematic method for testing
alternative hypotheses regarding interactions between variables, and allows
interactions to be ranked numerically in terms of relative importance. The
diameter, type and number of supports used were found to have the strongest
associations with locomotor repertoire, suggesting that these variables, rather
than height, contextual behaviour, or the age-sex category of the orangutan,
have been the most important influences on the evolution of orangutan locomotor
profiles.